Conservation & Threats
There is no information on specific threats to the Pyura sp and no species of ascidian is currently lists on the IUCN red list. This species does however live on coral reefs which are under threat from global warming, ocean acidification and the effects of coastal development (McKeown, 2010). One of the issues affecting the Southern Great Barrier Reef is the effect of sedimentation from dredging in Gladstone Harbour. The following is a study on the effects of sedimentation on the feeding rate and behaviours of the ascidian Pyura sp.
Introduction:
Increased sedimentation from terrestrial runoff and coastal alterations such as dredging pose a major threat to coral reef ecosystems and the species that inhabit them (Fabricius, 2005; Rogers, 1990). High levels of suspended sediment are known to affect the growth and condition of filter feeding organisms by clogging their feeding structures and reducing feeding time and efficiency (Nicholls et al., 2003). As a filter feeding organism living on the Great Barrier Reef, Pyura sp is likely to be threatened by increased suspended sediment. The amount of food that ascidians consume, and hence growth rate, is dictated by the total time spent feeding, the filtering capacity of the animal, and the amount of food in the water (Kott, 1989). Ascidians are known to close their siphons in response to suspended sediments and other disturbances, reducing the total time spent feeding (Kott, 1989). This study investigated the response of a species of ascidian from the genus Pyura to increased suspended sediment load. The hypothesis that less food would be filtered from the water column by ascidians subjected to suspended sediment than ascidians with no suspended sediment was tested.
Method:
Two adult ascidians of the same species from the genus Pyura were collected from the South-West reef crest of Heron Island. The ascidians were each placed in a container with 200mL seawater and 0.5mL of 6% dry weight 7-20μm algal solution. After 1 hour 100mg of 125μm sediment was added to the water of one ascidian, while the other ascidian was left as a control. Every 15 minutes the solution was mixed to resuspend any algae and sediment. The algae concentration was measured at time zero and every 15 minutes for two hours to determine the feeding rate of the ascidians. Chlorophyll a was extracted from a 1mL sample taken at each time point using the method from Newton (2002). Chlorophyll a concentration was measured using a spectrophotometer then used to calculate algae concentration.
Observations on the feeding activity noted by the opening and closing of the siphons were taken.
Results:
No change in algae concentration over the two hours was found in the water for either the control or treatment ascidian (figure 1). Feeding rate was measured as zero for both ascidians.
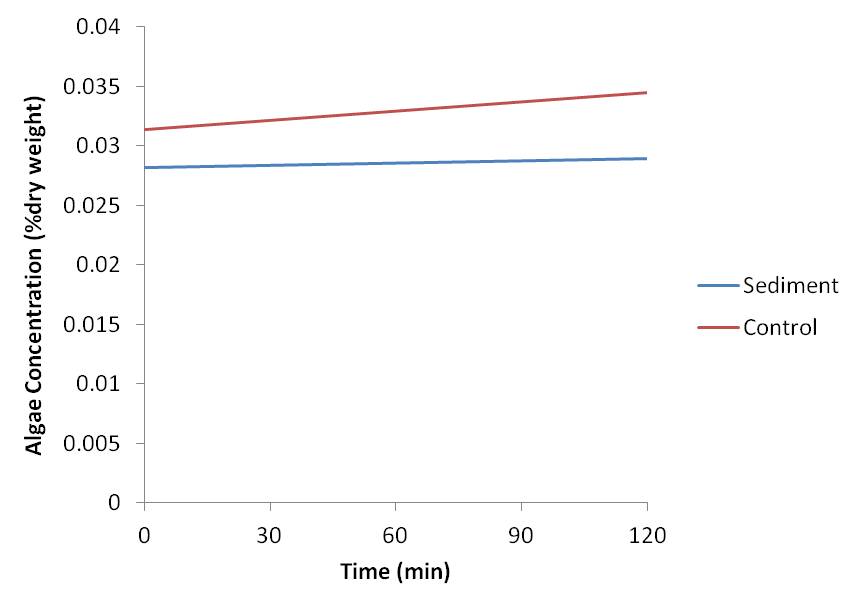
Figure 1. Change in algae concentration as a result of feeding in two Pyura sp with sediment added to solution after 60 mins (sediment) and without sediment added (control).
Both ascidians were observed to have their siphons open most of the time, indicating feeding. Feeding stopped sporadically for short periods in both ascidians, indicated by the closing of the siphons. The sediment treated ascidian closed its siphons and contracted its body immediately upon the addition of sediment (figure 2). Feeding commenced again within five minutes. Each time the water was mixed the sediment treated ascidian closed its siphons and contracted. The control ascidian showed no response to mixing and was never seen to contract its body to the same extent as the sediment ascidian
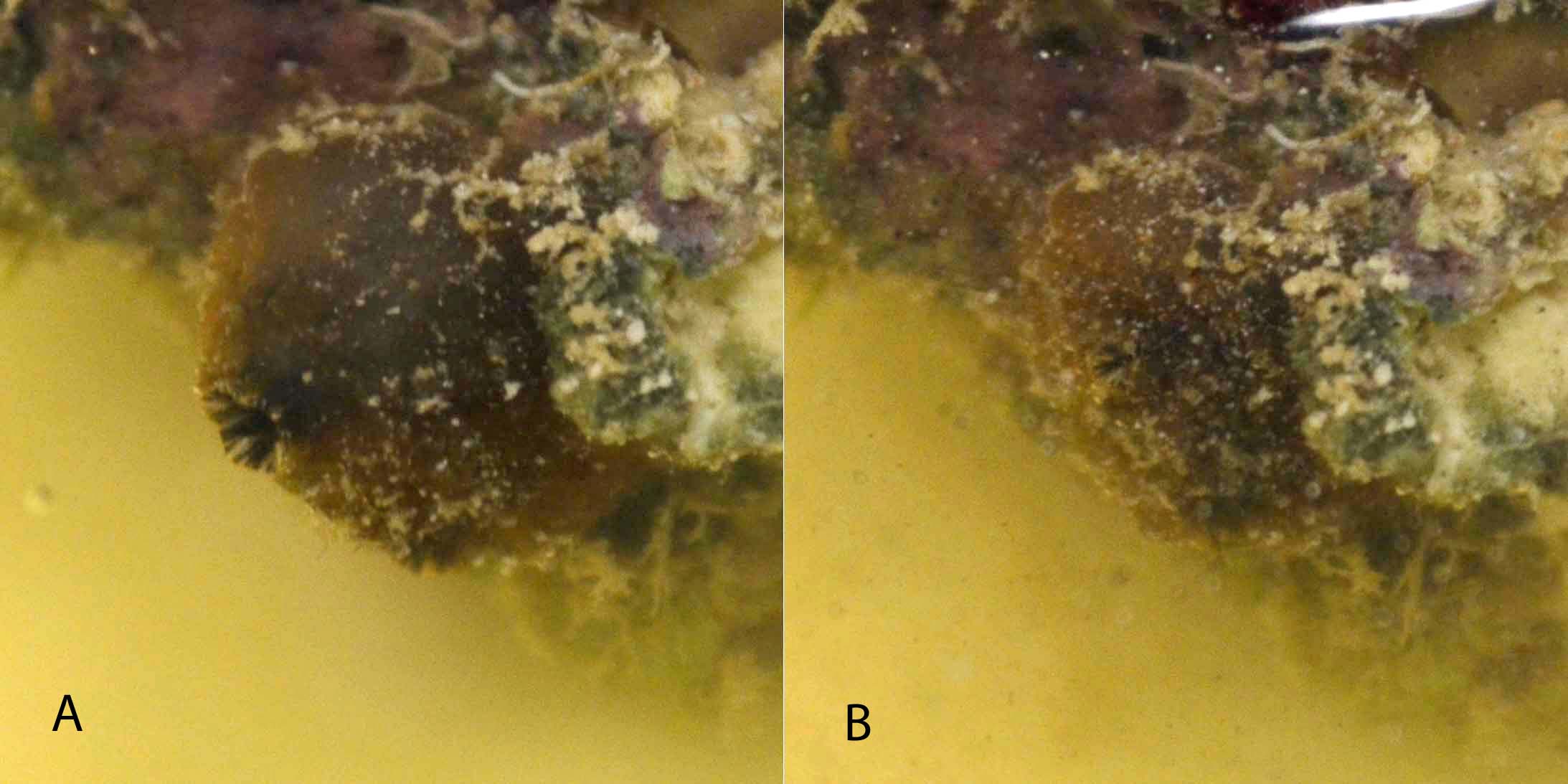
Figure 2. The sediment treated Pyura sp A observed feeding before the addition of sediment and B retracted after the addition of sediment.
Discussion:
No difference between the feeding rates of the ascidian subjected to suspended sediment and the ascidian with no suspended sediment was found as feeding rate for both was measured as zero. Both ascidians were observed with their siphons open during the majority of the two hour indicating they were feeding (Kott, 1989). It is possible that the volume of water the ascidians were placed in was too large and the algae concentration too high to measure a change in response to feeding in the ascidians. Feeding rate of the ascidians may have been low due to the still water compared to the currents present in the natural environment as increased flow of water increases the feeding capacity of ascidians (Kott, 1989).
Observations indicated a difference in behaviour between the sediment subjected ascidian and the control ascidian indicating an effect of sedimentation. The behaviours displayed by the ascidians fit with those found by Armsworthy et al. (2001) in their study of Halocynthia pyriformis, another stolidobranch ascidian. The closure of the siphons and retraction of the body of the ascidian in response to the addition or resuspension of sediment is likely to be the squirting response (Kott, 1989). When exposed to high sediment loads, H. pyriformis was found to close its siphons and squirt out up to a third of the water from the pharynx and atrium. This is done to reject any unwanted particles and reduce the risk of clogging (Armsworthy et al., 2001).
The results of this study suggest that this species of Pyura has behavioural mechanisms similar to that of H. pyriformis to mitigate the effects of suspended sediment on nutrient acquisition. Further studies need to be done to measure the effectiveness of these behaviours and test the difference in feeding rates of ascidians when subjected to suspended sediment and not.
|